--科学による価値探求の手法--
前書き
脳の研究
科学技術の分野において何の研究をするべきかという判断は、非常に難しい。研究そのものが1000年後においても有益と予想される、人生をかけるに値する真に意味のある研究である。
真理を探究する理学および社会への技術的応用を目指す工学の双方において、複雑化する科学技術の価値を宇宙・社会・生き物と関連付けて明確にすることは、重要なことであろう。この考察だけでも一つの大きな学問となるくらいであることは想像に難くない。例えば、科学哲学という学問分野がある。
生き物の中でも、人とのかかわりは、研究の価値を考えるうえで特に重要な要素となる。脳が人の思考の中枢と考えれば、人と社会の仲介をしているのは、脳である。人の研究、中でも脳の研究は、その解明の難しさとあいまって学問の中で重要な位置を占めている。

図1 シュレディンガーの猫
1.1 量子論
量子理論の不確定性原理や遺伝子に基づく進化論など、科学は価値体系に大きな示唆を与えている。不確定性原理は世界が機械仕掛けで運命が定まったものではなく、予測不能な可能性にあふれていることを示唆している。
不確定性原理によると、エネルギーと時間の積において、不確定さがあるため、その両方を正確に測ることはできない。不確定性原理と巨視的な現象の関係を一般の人に説明するうえでよく知られている例は、「シュレディンガーの猫」と呼ばれるものである。量子的な過程で、箱の中の猫が死んでいるのか生きているのか決まる場合、箱を開けてみるまで分からないことになる。マクロには、そうしたことはあり得ないが[1]、微視的な領域においては、こうしたことは頻繁に起こる。
もう一つは、カオス理論におけるバタフライ効果といわれているものである[2]。蝶が羽ばたいただけでも、長期的に見れば天候に影響を及ぼすことがあり得る。バタフライ効果も、この世の中が、予測不可能で不確定であることを示唆している。
ペンローズは、脳の働きの根源を量子効果が支配していると考えた[3]。これも、予測不可能な心理現象を示唆しているのであろか。私は原理的な部分では古典的な論理で説明できると考えている。その理由は、核スピンという量子計算の容易なメモリにおいても、ロジックを構成できるほどには、理論的には働かないからである[4]。量子計算をするには各スピンの独立性が高く、外からの擾乱を受けないことが必要である。核スピンは、スピン軌道相互作用によって、電子スピンの影響を受けるが、電子スピン同士の相互作用に比べるとその影響ははるかに小さい。液体中では、核スピンの独立性が高く、それを用いて量子計算が可能であるが[1,5]、固体中では、難しい。相互作用が固体中となる脳内では、核スピンの独立性を壊す、デコヒーレンスと呼ばれる効果が大きいことが分かっている[6]。脳内量子計算で、高密度なロジック回路を構成するのは、難しいと思われる。

図2 遺伝子と進化
1.2 遺伝子論
科学的な価値体系を決める一つの土台はダーウィンの進化論[7]、ネーゲリーの染色体発見、メンデルの遺伝の法則、ワトソン・クリックの遺伝子の二重螺旋発見に至る一連の研究によってもたらされた[8]。人の可能性には遺伝子という枠がはめられていることを示唆している。
遺伝子の概念から社会的考え方にもたらされる帰結の一つが、遺伝子が人間を操るという利己的な遺伝子論である[9]。

図3 コンピュータとメモリ
2.1 価値体系
フリードリヒ・ニーチェのニヒリズムと仏教は絶対的な価値体系から、一歩引いているという意味で共通した思想である。ニーチェは「絶対にこれが正しい」「絶対に美しい」といったものを否定した[10]。一方、仏教は、始祖である釈迦の後に長く続く、膨大な思索がある。その思索の一部は般若心経にまとめられている[11, 12, 13]。般若心経では遠い宇宙から地球を見ているかのように、人間の一切の恣意的な考え、価値を相対化している。
ニヒリズムや般若心経が示唆するように、すべての価値が相対的であるならば、脳が器としての存在を超えて、価値を創造する源の地位を持つ。脳の働きを解明することは価値基準の源を探求することそのものとなる。脳の働きと人間の行動との間を喜怒哀楽という言葉で表すことはできるが、細胞レベルで見て、喜怒哀楽が何かを示すのは難しい。脳の働きを詳細に解明するためには、細胞の形状やその変化を計測する技術が必要であり、この技術は、急速に進歩している。
2.2 脳と価値体系
量子論や遺伝子理論が、価値が形成される原理の説明であるのに対して、脳は直接的に価値に基づいた行動に影響する。外からの情報を処理し、体の各所を動かす行動原理を支配しているといえる。感受性の5感や、食欲・性欲である。これらは、コンピュータの起動時に必要なファームウェアやシステムファイル、プログラムとハードウェアの仲立ちをするオペレーティングシステム(OS)などに相当する。一方、その人の個性を形作る重要な因子が記憶である。記憶に基づいて、行動原理が発揮される。記憶は、一時記憶と長期記憶があるが、それぞれ、一時的な記憶装置であるメモリーの一種であるダイナミック・ランダム・アクセス・メモリ (Dynamic Random Access Memory:DRAM)と長期的なデータの保存に適したメモリーであるハードディスクや光記録媒体に当てはめられるだろう。コンピュータはDRAMを含む様々なICチップからできている。ICチップは、コンピュータの中に入っている集積回路である。縦横数mmから数cm、厚さ数mmの基板の上に多くのトランジスターが並んでいる。2022年時点で、ICチップは基本単位であるトランジスタを数百億個含むようになってきた[14]。
コンピュータの動作を決めるのはソフトウェアである。ソフトウェアがなければ、動かない。コンピュータにどんなソフトウェアが入っているかを知れば、それが、何を目的とし、どのような動作をするか知ることができる。ソフトウェアは複雑な動きをするが、基本原理は、足し算、引き算の命令を出すソフトウェアと、それに対応するハードウェアからなっている。脳ではそれほど単純な動きをしているとは思えないが、まずはこの対応を知るところから研究が始められると予測できる。
脳が価値に大きな影響を与えると考えれば、脳の研究は人文と科学をつなぐ可能性のある重要な研究対象であることが分かる。脳の働きについては膨大な研究があるけれども、重要なことが ほとんど分かっていない。計算機では、記憶の単位がトランジスタで作るメモリセル(記憶素子)であり、情報は01の2進数で記憶されている。これが計算機を構成する最も基本となる単位である。一方、脳について、その01に相当する情報がどのように記憶されているか不明である。そのため、記憶された情報がどのように処理されているかは、さらに曖昧にしかわからない[15, 16]。
記憶に関する最もわかりやすい脳の現象は、おばあさん細胞(Grandmother cell)である。サルがおばあさんの顔を見ると特定の細胞一つが強く反応する。あたかも、一つの細胞が一つの記憶に対応しているようである[17]。人間の脳を構成するニューロンの数は500 億から1000 億の間であり[18, 19]、そのうち100 億は大脳皮質に属している[20, 21]。トップレベルのスーパーコンピュータの場合、IC チップあたりのトランジスタ数は10 の9乗[22, 23]、コンピュータあたりのチップ数は10 の5乗 である[23]。すると、トランジスタ数は1,000,000 億(10 の14乗)になる。ニューロンの数に比べると桁違いに多い。しかし、人間のように様々なことをこなせるわけではなく、脳に比べて優れているとは思えない。一つの細胞が一つの記憶に対応していると考えたのでは、この優位性の説明がつかない。一つの可能性は、もっと複雑な、細胞と細胞のあいだのネットワークがあることである。例えば、細胞間の距離や細胞の形状が情報源となっていれば、情報量は増える。
おばあさん細胞モデルでかつ、フーリエ変換のようなニューロンの間隔からホログラムへの変換がなされているなら、強い信号だけでなくそれに伴う微弱な信号が重要となる。そう考えると、この微弱な信号を読み取る計測方法が重要である。ニューロンのパルス信号や、ニューロンの形状がパラメータとなり、信号はニューロンの形状で修飾を受けるのかもしれない。検討事項は、情報の鍵は強度なのか信号の間隔なのかという問題や、信号はどのように分岐するのかという問題となるであろう。
脳の現象論としては、神経情報の伝達経路や、記憶場所がどこかなどについて、膨大な研究がある。しかし、記憶メカニズムは分かっていない。そのメカニズムが不確定現象である量子論的か、それとも機械仕掛けの時計のように古典的かが論じられている段階である[3]。
モデルも様々であり、各モデルが重要視する観点も異なる。連想記憶はホログラフィックメモリで説明できるというGaborの説は[24]、連想が得意な人類の脳に着目したものである。他に、心が脳だけでなく内臓の影響も受けているという説がある[25]。さらに、ニューロンネットワーク以外に賦活をつかさどる機能構造として小脳チップや大脳チップがあり、記憶の根源となっている説がある[26, 27]。また、電気的なモデルで計算して、脳の働きを説明しようとする方向もある。例えば、脳の各部位の信号のやり取りで、学習過程を説明し、人間が判断をする仕組みを明らかにしようという試みである[28, 29]。
脳は、おおまかに大脳小脳に分けられる。脳の器官のうち海馬が記憶にかかわっているとされており、これは大脳の下に位置する。この近くには、視床があり、大脳に至る視覚情報の伝達経路となっている。形状情報は、フーリエ変換され三角や四角などの情報に分別される[30, 31, 32]。このように、記憶の情報の経路や大まかな保存場所については研究が進んでいる[33]。
一方で、画像記憶がどこにどのように保存され、それがどのように呼び覚まされるのか分かっていない[34]。求められているのは、「包括的な理論的枠組み、すなわち心の事象が、脳の活動パターンにいかに関連するかを余すところなく説明する、一連の架橋的な法則を見出すことである」[35]。その前提となる記憶の痕跡(Engram)はどこに残っているのかということも明確でない。例えば、おばあさんの顔など、特定の画像情報に単一の細胞が強く反応する。しかし、細胞レベルで見て、情報がどのように表現され、どのような形態で保存されているかはわかっていない。そして、問題は、何を明らかにすればこの手掛かりが得られるかである。それは、何を計測すればよいかという、現実問題につながる。
これまで膨大な研究がなされているにもかかわらず、記憶の痕跡がどこにあるかつかめていない状況は、新しい研究手法を取り入れる必要性があることを示唆している。
器に過ぎない脳が、価値の源として寄与している可能性について述べてきたが、その解明は、価値と結びつけるところまでは来ていない。脳の解明には様々な、知識が必要である。脳の成長過程、記憶や信号伝達のメカニズム、そのメカニズムに基づく脳のモデルが挙げられる。他の臓器との結びつきの有無も重要である。人間の行動原理については心理学で研究されているが、最終的には、脳のミクロな機能まで統合して理解することで、価値と結びつけた議論が可能となるであろう[36]。
重要なことは、意味や価値が、神経の情報伝達における記憶や信号処理とどう結びついているかである。意味や価値という抽象的な概念が、信号レベルでどのように表現されているかという問題である。例えば、恒常的な反復過程において、異なる応答が出てくることが、何らかの意味を示しているのかもしれない。あるいは、特異的な入力パターンで応答が変化することが重要なのかもしれない。
5感に対する外からの信号が、常に同じである場合を考える。例えば、窓や物がなく壁だけの部屋に監禁されて、かつ目を閉じていた場合である。外からの信号がなくても、すでに多くの記憶が形成された成人であれば、何かを考え、創造することができる。脳の内部だけで意味や価値を創出することができるのであれば、外部信号が定常的であっても、信号の出力内容は自発的に変化すると考えられる。
同じ入力であっても、時系列では、異なる応答を示すはずである。ニューロンの繋ぎ変えや修飾によるものかもしれないし、信号の再現性によるものかもしれない。記憶がどのように蓄積され、価値観が形成されていくかが、脳と社会を結びつけつ重要なカギであろう。
心理学は人の行動原理について多くの考察を行ってきた。脳というブラックボックスについて、様々な症例をもとにその特性を検討してきた。今後は、もっと物理的な脳に根差した証拠と心理学的な事象を対応させることでより理解が進むだろう。例えば、脳の遺伝改変マウスを使った実験は、脳の機能と特定の遺伝子との関係を明らかにしている[37]。さらに、価値と脳の関係を結びつけるためには、特定の記憶がどのようにして、好ましいものとされ、行動に結び付くかを明らかにしていくこととなる。
計測技術は急速に進歩し、多くの脳のモデルが提案されているが、それでも、まだ脳の解明に寄与するレベルに到達していない。例えば、計測のスピードや分解能が不十分とされる。脳のモデルは、量子的かどうかという原理的な点についてまだ議論されている段階である。
脳の部位である大脳・小脳・前頭葉・側頭葉などの区分について、ミクロな構造が計測され、各部位の大まかな役割が明らかになってきている。計測端子としての電極や特定個所のマーカーとなる蛍光を用いた計測は、基本単位であるニューロンについて、分子レベルで、その機能が明らかにしてきた。
このように脳の研究には多くの努力が積み重ねられてきた。しかしながら、様々な側面の知見のうち、脳のメカニズムに関する部分は、最も解明が、進んでいないと思われる。ブルーレイの光メモリは一つのビットを単位として記憶している。一点ごとに、反射率の高低で1か0かを判別している。脳でこれに対応する、記憶の痕跡が何かすらわかっていない。つまり、記憶の原理が明確には分かっていない。例えば、おばあさんの顔を覚えるときに、コンピュータでは、1と0の情報にまで分解してIC回路に記憶するが、これが、脳ではどのような情報として保存されているか不明である。MRI(Magnetic Resonance Imaging )による脳の断面像やニューロンの電極による電位計測は、膨大な情報をもたらしてきた。さらに、空間・時間の分解能や感度、侵入深さ、元素の区別、分子構造変化の検出など多くの面で計測技術の向上を図ることが求められている。特に、脳の基本単位であるニューロンに着目している。注目点は、隣接するニューロンへの信号の分岐と、各信号の短い時間内での強度の変調である。これを高速・高感度・高精度・非破壊・非接触で計測すれば新しい情報が得られる可能性が高い。
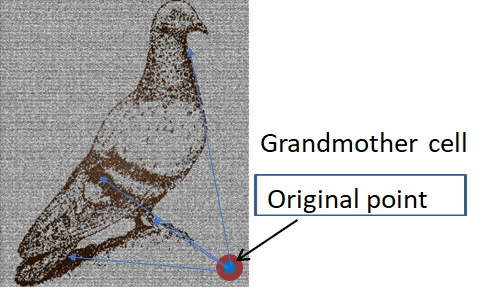 図4 おばあさん細胞のニューロンと鳩の像 [16]
図4 おばあさん細胞のニューロンと鳩の像 [16]記憶の痕跡はニューロン形状のわずかな変化であるといわれている[38]。しかし、一つの細胞でそのような複雑な顔を記憶できるのだろうか。
ホログラムという概念がある。ホログラムメモリでは、情報が面に記録され、例えば、入射角度を変えることで、複数の情報を読み取ることができる。光では、波面が維持されることで干渉が起こることを利用している。脳の信号がミエリン鞘の中で、干渉性が維持されるのであれば、同様の干渉効果が起きてもおかしくはない。入力情報が明確な視覚が最も解析しやすいと思われる。視覚経路の研究は進んでいるが、記憶とどう結びついているかは明らかでない[39]。
ホログラムメモリは記憶領域が広がって非局在化しているが、局在している記憶モデルもある。イメージがそのままの形で保存されているというモデルである。波面がイメージおよび基準となる強い散乱点で散乱されるとき、その散乱パターンをフーリエ変換することで元の像が復元できる。
この光散乱の計測において、散乱パターンのコントラストは、散乱体の直前に集光点があるときに最も高くなる。散乱体が二つあれば、二つの散乱体の距離でパターンが決まる。この散乱体間の距離を情報とすることで記憶モデルを構築できる。このモデルが神経細胞の記憶に適用できるならば、おばあさん細胞を複数ある散乱体のなかで散乱強度の大きい中心となる柱と考えて、おばあさん細胞とおばあさんの情報に対応する細胞を含む領域に、位相のそろった電気信号の波が来た時に最もコントラストが高くなるのではないか。
これは、その細胞を起点として、他の細胞との距離から情報を形成しているというものである。もしそうならば、観測できているのは、強い信号だけで、真の情報は他の細胞に流れた微弱な信号の中に含まれると考えられる。
おばあさん細胞から微弱な信号計測の必要性が推測できた。微弱な信号が記憶に影響を与えるとすると、ニューロン一つ一つの信号を感度高く計測する必要がある。信号はmsオーダーの間隔で並ぶスパイク系列であり、高速性と高分解能を両立することが重要。近年の高感度ハイスピードカメラは1フレーム1μs近い計測速度を、高い感度とダイナミックレンジで実現している。
注釈)たとえば、Photron製 ハイスピードカメラ FASTCAM Mini AXは、
640×480ピクセルで20,000コマ/秒、という撮影速度を実現したハイスピード
カメラである。モノクロ12bitという高精細画像に加え、モノクロISO40,000と
いう超高感度性能を実現している[40]。
脳はニューロンが格子のように並んだ神経ネットワークであるが、従来の研究の多くが、せいぜい単シナプス伝達にとどまっている[41]。現実には、単シナプス伝達も正確に解析できていない。単シナプスの解析には、電極を神経細胞に差して、電位の変化を計測する方法が主流であった。しかし、この方法では、電極が信号に影響を与える可能性がある。非接触で計測する手法としては、光計測が有力である。ホログラフィーによる3次元計測や、二光子吸収による3次元計測で高速で高分解能な手法の検討が進められている。
複数のニューロンの信号を光学的に計測する手法としては、ホログラフィーや二光子吸収以外にも、光波散乱計測を利用するものがある[15, 42]。
ホログラムは空間的な集光・検出位置を容易に制御できる点で優れている。二光子吸収は脳の深いところまで計測できる点に特徴がある。一方、光波散乱計測は分解能と計測の高速性で優れている[43]。
脳内の信号の計測方法は、そのとらえる対象によって計測方法が異なる。脳の計測対象は、最表面、数ミリの深さ、脳全体で区分できる。さらに、分解能や計測スピードが重要な因子である。計測方法では、非接触、接触、破壊計測で区分される。
脳全体を計測できる手法はX線CTやMRIである。これらの手法は、脳全体の構造を非侵襲に計測することができる。特に、MRIは、構造だけでなく、分子の運動性もとらえることができる。その面内分解能(in-plane resolution)は、14μmが可能である。さらに、脳の特定の部位、例えば、脳梁(corpus callosum)にある神経の束について、その向きと共に直径を判別できる。例えば、2~4μmにあるものと10μmのものを見分けることができる[44]。
脳の表面の場合、例えば、原子間力顕微鏡(Atomic Force Microscope: AFM)および走査イオン伝導顕微鏡(SICM)は、探針を用い細胞の表面をなぞることで、その立体形状をナノメートルの精度で計測できる[45]。SICM は、神経細胞と探針との間の距離の変化により生じるイオン電流の変化を利用することで、試料に触れることなく表面形状や帯電を画像化できる[46]。計測スピードは25 μm^ 2/min と遅いが、非破壊計測における分解能は、他の手法と比較して最も高い[47]。
電極で神経細胞の電位を図る方法が、接触方式ではあるが、脳の情報伝達については最も多くの成果を上げている。その一つ、パッチクランプ法は、イオンチャネルやトランスポーターを介したイオンの挙動を記録する方法である。神経伝達物質トランスポーターは、神経終末より遊離された神経伝達物質を再取り込みすることによりその神経伝達を終結させる[48].
脳の価値基準を担う器としての機能を評価する重要なポイントは記憶がどのように保存されているかであろう。外界と相互作用の結果、記憶に基づいて判断することで、人の行動が決まる。
これまで電極による手法が使われてきた。脳の内部まで計測可能であることや、特定のニューロンの活動を計測できることがその理由である。有力な方法であるが、脳の記憶の痕跡をとらえるには至っていない。この理由は、接触式であるために、その計測自身が神経に影響を与えてしまうことが一つの原因と考えられる。神経の微小な電気信号をとらえるには向いていない。
この電気信号を非接触で計測する方法の代表格は、光計測である。
9.1 微細形状の光計測
光学顕微鏡で微小試料の形や色を正確に見たいというのは多くの人が共通して持つ希望である。現実には、サイズや屈折率が複雑に見え方に影響するため、その希望がかなえられるケースは、限られている。近年の光の伝播を計算する技術の進歩は、この問題を解決する手がかりを与えつつある。形状やサイズが色に影響を与えるよく知られた例は、シャボン玉の虹やモルフォ蝶の翅である。通常、色は複素屈折率の虚部に大きな影響を受ける。しかし、構造によっては、複素屈折率から想定される色とは全く異なる色になる。
一方、形状・サイズは複素屈折率によって、見え方が異なる。膜厚の計測において、屈折率の情報が不可欠であることがよく知られている。未知の微小物の形状や複素屈折率を知ることは、計測された形状・サイズ・色の情報から、これらのパラメータを推定することに他ならない。
色が構造によって影響を受けるのは、サイズが共鳴領域といわれる波長サイズの領域で顕著である。試料が粒子の場合、共鳴領域の粒子径は、波長の1から10倍程度である。線形近似では、光は試料面で、屈曲ないし直進して、光束が維持される。しかし、この粒子については、界面で光束が広がってしまい、この広がりは波長によって変化する。そのため、反射率や散乱パターンは、波長ごとに異なる[49, 50, 51]。
9.2 光計測の分類と光波散乱計測
光を利用して小さいものを見る原理(顕微鏡の原理)は、その近似の程度から数学的に大まかに3つに分けられる。一つは光が直進することを利用して、光線追跡を行う方法である。線形近似である。試料内を光は直進し、試料の像はそのまま投影される。医療用に広く使われる、X線CTではこの投影像から元の3次元像を再構成する。もう一つは、レンズの結像を利用する方法である。光学の原理としては、レンズのフーリエ変換作用を利用している[52, 53]。そして、フーリエ変換ができるのは、フラウンホーファー近似あるいは同様の近似である一次のボルン近似を用いたときに限られる。フラウンホーファー近似は、試料サイズに比べて十分遠方で計測されるときに成立する[54]。そのため、近距離の光の伝搬の考慮が必要となる試料内での多重反射や多重散乱は考慮できない。3次元形状を計測するための主要な手法の一つであるホログラフィも同じ近似に基づいており、試料表面で反射された光は自由に伝搬すると考える。3番目が、試料の厚みや屈折率を厳密に考慮して、散乱パターンから像を推定する手法である。光波散乱計測と呼ばれている。元の像を求める過程は試行錯誤であり、単純な形状でなければ適用が難しい。しかし、解析がうまくいけば、他の二つの原理と比較して計測対象サイズを二桁小さくでき、二桁正確度が向上する。3次元の高分解能・高速計測の手法として、光波散乱計測(スキャトロメトリー)が有効である。従来の主な高分解能3次元計測手法はレンズ結像、ホログラフィ、位相再生法である。ホログラフィや位相再生法は3次元像を高速に得るのに適しているが、フーリエ変換を用いることを前提としているので、フラウンホーファー近似の仮定が入る。このため、散乱や吸収が大きい場合、これらの手法では、計測対象のサイズが小さくても100波長程度にとどまっており、正確度も10波長程度である[55]。3次元像を高分解能に計測する手法としては2光子吸収あるいは、一分子の蛍光を時間分解で高感度計測する手法が知られているが、これらも、レンズの結像を利用しているので、フラウンホーファー近似の範疇に入り、吸収や散乱が大きい場合には、同様の性能となる。光波散乱計測は、光学の原理であるマクスウェルの方程式を厳密に数値計算で評価する。このため、3次元形状計測において、フーリエ変換を利用した解析に比べ散乱や吸収が大きい場合、正確度が二桁向上する[56]。
光波散乱計測は計測速度が速い点もメリットである。光波散乱計測では、数波長~数十波長程度の領域の高分解能3次元計測をワンショットで行えるため、蛍光の減衰時間が律速となる超解像計測や、一点ごとの計測となる二光子吸収計測と比較して、計測速度が速い。
可視光・紫外や極短紫外・軟X線では、しばしば、媒質と試料との屈折率や消光係数の差が大きく散乱の影響を受けやすいので、光波散乱計測は特に有用である。10波長程度の試料サイズでは、レンズ結像やホログラフィの基礎原理であるフラウンホーファー近似が成立しない。試料が薄い場合には、回折散乱効果が小さいので、この近似で解析可能であるが、試料が厚くなると、難しくなる[57]。
光波散乱計測の欠点は解析が難しく、元の像が一意には定まらないことである。他方、線形近似やフラウンホーファー近似では、元の像と散乱パターンを1対1で対応させることができる。数学的な解は確実に求まりそのスピードも速い。
光波散乱計測で、散乱パターンから元の像を求めることを逆問題を解くといっている。逆問題を解く手法として、深層学習や最小二乗法が取り入れられている。現状では、柱状で、柱の断面が矩形や三角、楕円などである単純な形状に限られている。
光波散乱計測は、可視光・紫外での計測のように、波長に制限がある場合や、X線による有機物の計測のように、短波長での試料の破壊が問題となる場合に力を発揮する。scatterometryは、可視光・紫外での計測のように、波長に制限がある場合や、X線による有機物の計測のように、短波長での試料の破壊が問題となる場合に力を発揮する。光波散乱計測を使えば、同じ分解能であれば波長を100倍長くできる。この効果を考慮して、試料の損傷を同じように抑えた場合、分解能10倍強向上する。
光計測は非破壊・非接触であり、高速な計測が可能である。 電子線を含め、一般にエネルギーが高い(波長が短い)方が分解能が上がる。一方で、エネルギーが高い方が計測光による試料の破壊が起こりやすい。[58]。これは、生体など有機物の場合には特に問題となる。
このトレードオフの結果、分解能と試料破壊を両立できる限界が、従来は、波長10 nm程度になっていました。
9.3 コヒーレント光散乱とその限界
レーザーの光散乱は完全なコヒーレント光と考えることができ、数学的に簡潔に記述できる。一方、ハロゲンランプなどの白色光では、試料サイズや波長に応じて、インコヒーレンスの程度を考慮した扱いが必要となる。コヒーレント光散乱は計算と対応させることが容易で、実験結果の解析が可能であるという意味で、有効な手法である。コヒーレント光がもののサイズを計測するのに適している理由は、部分コヒーレント光の特性がインコーレンスの度合いが不確定であり、その度合いに応じた計算が複雑であることによる。
コヒーレント光散乱は、原理的には光学的な計算手法により散乱パターンを正確に予測できる。微細試料の特性は屈折率の実部と虚部・形状・縦の長さ・横の長さ等多岐にわたるが、入射角度や波長を変え、透過・反射の散乱角度分布および反射率・透過率を計測することで実験データも多く計測できるので、おおまかな情報は得られる。
実際に試料を計測する段になると、コヒーレント光が試料内部でインコヒーレント光になってしまうという問題に直面する。内部での散乱体が小さく、散乱パターンへの影響が小さい場合にもこの問題が発生する。厳密な光散乱の計算は、コヒーレント光を仮定しているので、誤差が大きくなる。
未知試料の内部屈折率分布が正確に分かっていれば、計算可能であるが、微小散乱体の散乱強度は小さく、数も無数にあるものなので、分布を計測するのは難しい。この場合、コヒーレンシーを一つの指標とすることで対応が可能である。試料内部で一律にコヒーレンシーが低下すると仮定する。大きな近似であるが、全く何も考慮しない場合に比べて、実験との一致はよくなる[59]。
光波散乱計測は、近似をしないという長所があるが、その代わりに、散乱パターンから元の形状を逆フーリエ変換等を用いて直接求めることができないという欠点がある。この手法が使えるのは、元の形状が単純であるか、おおよその形状が既知である場合に限られる。神経細胞のように、おおよその形状が分かっており、一つの断面形状だけが分かればよい場合に適した解析手法である。
9.4 神経の高速・高分解能な3次元光計測
光を利用して小さいものを見る原理(顕微鏡の原理)は、前述のように、1.線形近似と、2.フラウンホーファー近似、3.厳密解を解く手法(光波散乱計測)に分けられる。神経の信号計測では、2番目の手法が、蛍光と併用して使われることが多い。蛍光体からの発光は、一分子の発光だけを時間分解でとらえることで、空間分解能の大幅な向上をもたらした。STochastic Optical Reconstruction Microscopy (STORM)である。この方法は、発光の頻度が多くなければ、時間分解能を上げることができず、励起光の強度、あるいは、試料の耐えられる集光強度にも限界があるため、計測速度と分解能の両立に限界があった。3番目の光波散乱計測は、蛍光標識を必要としないだけでなく、時間分解能の大幅な向上ができる可能性を持っている。
9.5 光波散乱計測による汎用的な光計測装置(高確度顕微鏡)
実際の試料はフラウンホーファー近似で表現できる像とはしばしば大きく異なる。吸収や散乱の影響が大きい可視光や軟X線では、試料内での吸収や多重散乱が強まり、このかい離は顕著である。このような場合、光波散乱計測(scatterometry)は、レンズイメージングと比較して、単純な形状の3次元測定で解像度を2桁程度向上させることができる。また、ミリ秒以下の時間分解能で測定することが可能である。これは、非破壊、非接触、簡便な測定方法である。ニューロンの信号伝達に係る屈折率分布の変化の高解像度かつ高速な検出が可能である[60, 61]。上述の光学計算技術の最近の進歩により、形状やサイズを定量的に分析することが可能になった。断面の屈折率分布と遠方での散乱パターンの関係を正確に計算できれば、屈折率分布を明らかにすることができる。散乱光の角度分布を計算する方法として、ここでは厳密結合波解析(RCWA:Rigorous Coupled-Wave Analysis)を使用する。この理由は、内部構造を容易に考慮でき、遠方界の散乱パターンを直接計算できるからである。 RCWAは周期構造のみを扱うが、計算方法を工夫することで孤立系にも適用できる[62]。また、特定の構造を選択するには計測個所への集光が必要であり、集光点とサンプルの位置関係の設定は再現性に大きく影響するが、散乱パターンのコントラストを調整して再現性を確保することができる[63]。
2番目の方法と3番目の方法を組み合わせることで、リアルタイムな結像と高分解能な計測の両方を実現することができる。猫のゴルジ染色大脳とヒトのワイガート染色小脳の標本における ニューロンの回折パターンを可視光による光波散乱計測例がある[43]。
9.6 光学の非線形性や時間差計測の利用
二光子吸収やラマン散乱における、光吸収や散乱の非線形性を利用した計測が使われるようになってきた。これらの手法は、非線形性を用いない計測と比較して、空間分解能を上げることができる。回折限界は分解能を規定しているが、これらはこの回折限界を超える分解能を得る方法である。これらの手法の結像はフーリエ光学の枠組みのなかで検討が進んできた。しかし、非線形性を利用することで計測点一点ごとについて、分解能を上げることができる。例えば、蛍光体が2光子を吸収する過程は、光の強度が強いところに限られる。これを利用して、一光子吸収のレンズの集光範囲より、狭い範囲に限定して、計測することが可能となる。二光子吸収では、入射光の波長は計測波長の倍である。これを利用して、生体の吸収や散乱が小さい波長を選択し、試料の奥深いところまで計測することが可能となる[64]。問題点は、3次元形状の取得には、多数の点を取得する必要があり、時間分解能が悪くなりがちなことである。
この他に、非線形ラマン散乱という手法がある[65]。非線形ラマンは、時間分解能・空間分解能ともに、線形ラマンより優れており、波長を選択することで細胞組織の深いところまで計測できる。[66, 67]。誘電率は、電場の強度によって変化する。この強度変化が大きいところだけを観測できれば、入射光の集光径よりも小さい部分を観測できる可能性がある[65]。ラマン散乱は、一般に感度が小さく計測時間がかかる。この問題を解消するために、二波長の入射光のエネルギー差と試料分子のラマンの振動モードを共鳴させることで信号強度を増幅させる方法が開発されている。
もうひとつは、STORMのように、時間分解を利用する方法である。一光子の蛍光の発光は、レンズの分解能を規定する回折限界の広がりを持っている。しかし、その中心を正確に評価できれば、実質的に高い分解能が実現できる。
非線形性や時間差計測を利用する手法の問題点は、時間分解能を上げるのが難しいことである。レーザーの強度が強ければ時間分解能が上がるが試料が壊れやすくなる。
論文リスト
変更履歴
- 2023.9.9 Version 1 2nd
- 2023.7.19 SSL certificate obtained 1st